
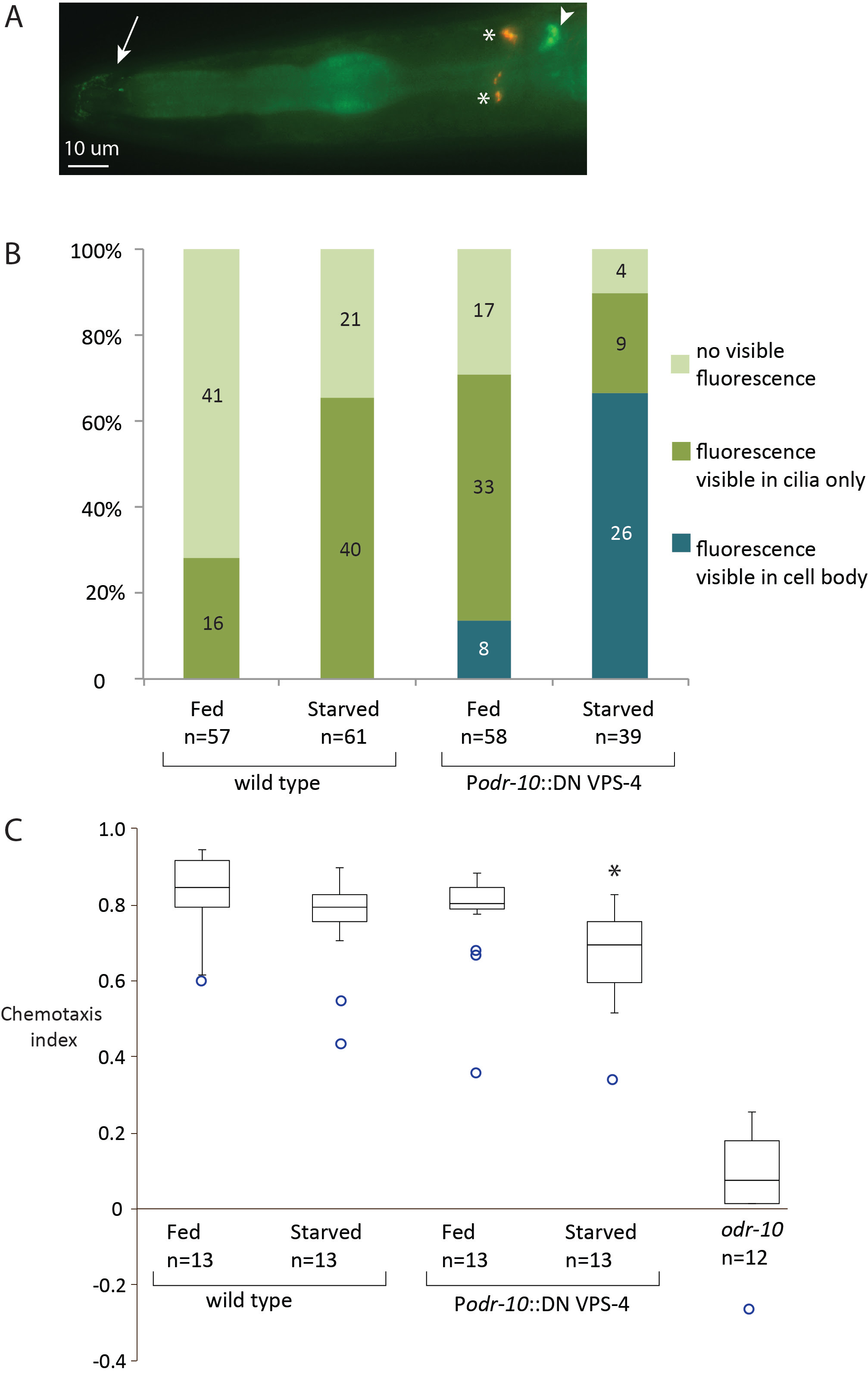
Description
Accumulation of ODR-10 depends on the sex and fed state of C. elegans (Ryan et al. 2014), though the precise mechanism of its regulation is not known. We tested whether the ESCRT pathway is required to maintain normal accumulation and distribution of ODR-10::GFP in AWA neurons under well-fed and starved conditions. The conserved AAA ATPase VPS-4 is required as the final step of the ESCRT pathway, to dissociate the machinery as cargo is moved from endosomes to multivesicular bodies (Babst et al. 2011, Kim et al. 2011). Mutation of Glutamate 219 to Glutamine (E219Q) in VPS-4 results in a dominant negative enzyme (VPS-4 DN). VPS-4 DN disrupts normal accumulation of the glutamate receptor, GLR-1, in a cell autonomous manner (Chun et al. 2008).
We investigated a potential role for VPS-4 in ODR-10 regulation using fluorescence microscopy and an attraction chemotaxis assay. We expressed VPS-4 DN under the odr-10 promoter in animals expressing ODR-10::GFP (Figure 1A). Using live-imaging (200X (total) magnification), ODR-10::GFP was not visible in over half of the wild type, well-fed animals. Using this baseline, we measured changes in ODR-10::GFP (abundance and localization) and found that VPS-4 DN increased the frequency with which we observed animals with aberrant ODR-10 accumulation (Figure 1B). In 8 animals (out of 58) there was observable fluorescence in the cell bodies of AWA neurons (1 with fluorescence exclusively in the cell body). Starvation exacerbated this effect: 26 out of 39 animals had fluorescence in the cell bodies of AWA neurons (7 with fluorescence exclusively in the cell body). In contrast to previously published data that relied on different imaging parameters (Ryan, et al., 2014) we also observed that starvation of animals led to increased frequency of visible fluorescence in the cilia of AWA in WT animals (Figure 1B). Expression of VPS-4 DN had no effect on the ability of animals to react to the chemoattractant diacetyl under well-fed conditions (Figure 1C). However, VPS-4 DN expression in the AWA neuron resulted in significantly lower chemotaxis indices, compared to wild type, fed animals (Figure 1C, asterisk).
Starvation of animals has a subtle effect on the abundance of ODR-10 that accumulates in the cilia of AWA neurons, but that accumulation does not affect the efficiency of chemotaxis in wild type animals (Figure 1B and C). However, when food stress was paired with a disruption of the ESCRT pathway the aberrant accumulation of ODR-10::GFP correlated with a modest but significant decrease in chemotaxis index (Figure 1C). It is still unknown whether ODR-10 is a direct target of the ESCRT machinery or other ubiquitin-related pathways. These data could also represent indirect effects due to changes in availability or localization of cofactors that are required for normal receptor processing and/or localization within the AWA neuron.
Methods
Request a detailed protocolAnimals were reared on lawns of OP50 on NGM plates according to established protocols. Well-fed, uncrowded young adult hermaphrodites were used for both imaging and chemotaxis assays. For assays using “starved” animals, L4 larvae were moved to NGM plates lacking bacteria 18-24 hours prior to assay. For fluorescence microscopy, young adult hermaphrodites were placed in 3µl M9 on coverslip, which was then inverted onto 3µl of solidified 2% agarose with 0.5% Levamisole (MP Biomedicals, Inc.) on the microscope slide. Animals were imaged on a Leica DMi6000 inverted microscope equipped with a Leica DFC3000G cooled CCD camera and GFP filter, at 200X total magnification (20X objective, NA 0.4). Animals were imaged using 150 ms exposures and gain of 10. Animals were described as having “no fluorescence” if no cells could be detected using these imaging parameters. Chemotaxis was assayed as previously published (Sengupta et al. 1996) using young adult hermaphrodites, 0.1% (v/v) diacetyl in ethanol was used as the attractant and ethanol as a negative control, with 0.02% (w/v) sodium azide as a paralytic at the site of the chemoattractant and control. Animals used for chemotaxis assays did not express ODR-10::GFP.
Reagents
Strains: N2 (Bristol)
CX3344 kyIs53 [Podr-10::ODR-10::GFP]
CLD5 dahEx5 [Podr-10::VPS-4 DN (E219Q); Pttx-3::dsRed]; kyIs53
CLD17 dahEx5; kyIs53
CX32 odr-10(ky32)
References
Funding
This work was supported by Research and Creative Opportunities for Undergraduates grants awarded to Ellen Zocher and Nelson Ruth by Western Washington University.
Reviewed By
Jay PieczynskiHistory
Received: July 23, 2018Accepted: July 31, 2018
Published: August 2, 2018
Copyright
© 2018 by the authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0) License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.Citation
Zocher, E; Ruth, N; Dahlberg, C (2018). Dominant-negative VPS-4 disrupts ODR-10::GFP distribution but has limited effects on chemotaxis. microPublication Biology. 10.17912/gnyw-v322.Download: RIS BibTeX
