Department of Biology, Hopkins Marine Station of Stanford University, Pacific Grove, CA 93950

Description
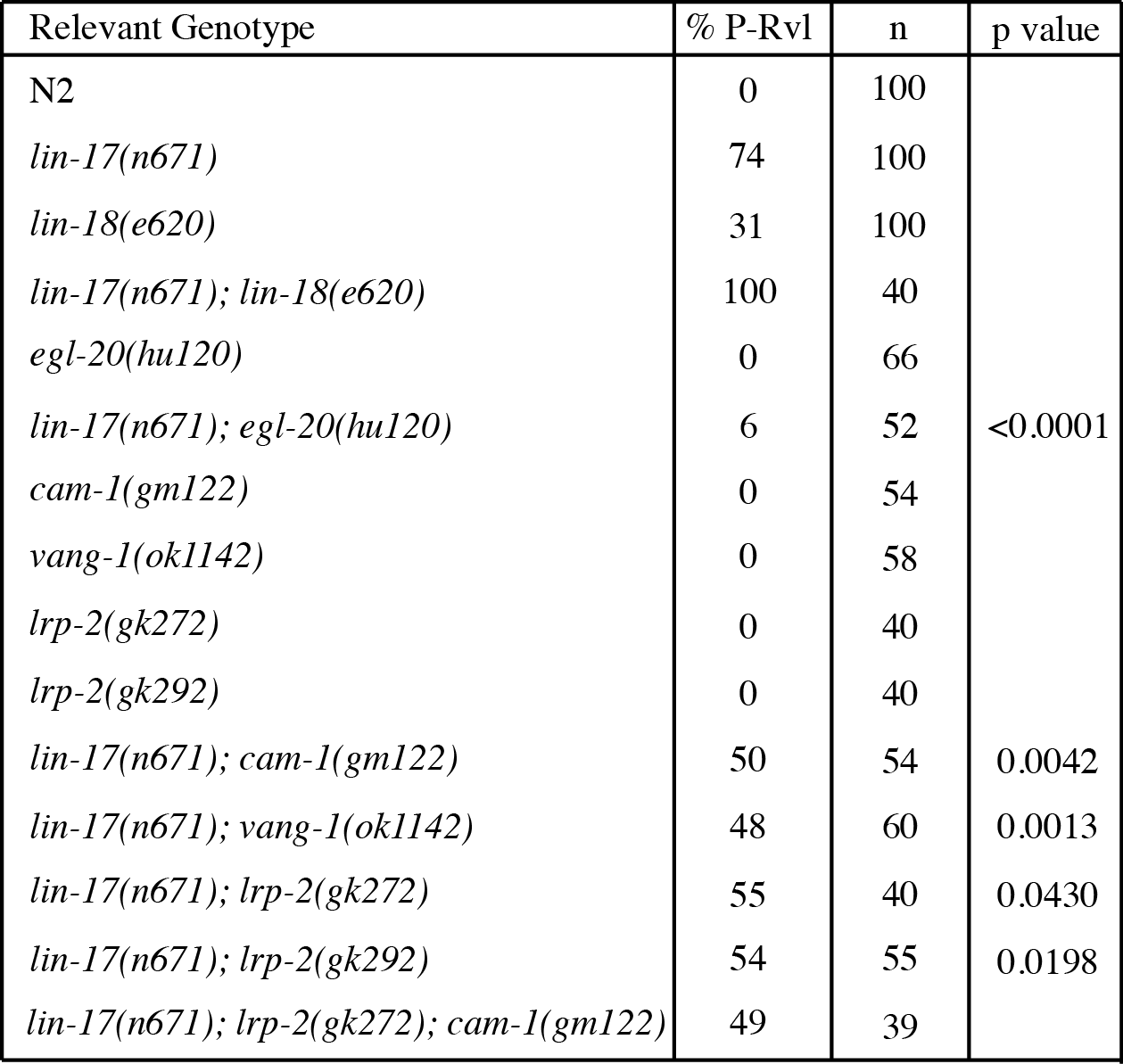
The C. elegans vulva is formed from divisions of three vulval precursor cells (VPCs) – P5.p, P6.p, and P7.p – arranged along the anteroposterior axis in the ventral epithelium (Sulston and Horvitz, 1977). Previous analyses show the orientation of P5.p and P7.p descendants is determined by the interaction of multiple Wnt signals. Specifically, in the absence of all Wnts, the VPCs display a randomized orientation, which is likely the default (Green et al., 2008; Minor et al. 2013). Two separate Wnts from the anchor cell, LIN-44 and MOM-2 acting through receptors LIN-17/Frizzled and LIN-18/Ryk, respectively, regulate P7.p orientation (Ferguson et al., 1987; Sternberg and Horvitz, 1988; Sawa et al., 1996; Inoue et al., 2004; Gleason et al., 2006). In the absence of these signals the orientation of the progeny of P7.p mimic those of P5.p and face toward the posterior of the worm, a phenotype referred to as posterior-reversed vulval lineage (P-Rvl). This posterior orientation is dependent on the instructive signal EGL-20, a Wnt expressed in the tail acting through CAM-1/ROR and VANG-1/Van Gogh, and is referred to as “ground polarity” (Green et al., 2008).
Here we examine the role of a low-density lipoprotein receptor, lrp-2, and its role in controlling the orientation of P7.p daughter cells. To investigate this interaction double mutants were constructed with both alleles of lrp-2 and lin-17(n671) (Table 1). Much like cam-1(gm122) and vang-1(ok1142), both alleles of lrp-2 suppress the lin-17(n671) phenotype from 74 to approximately 50% P-Rvl leading us to hypothesize that lrp-2 functions in the same pathway as cam-1 and vang-1. Furthering this hypothesis we have shown that, like cam-1 and vang-1, lrp-2 controls the localization of SYS-1/b-catenin (Minor and Sternberg, 2019). To ensure that this phenotype was a result of loss of lrp-2 function as opposed to background effects we injected a fosmid (WRM0617cA02) containing the full-length sequence of lrp-2 and found that it does rescue the double mutant phenotype of lin-17(n671); lrp-2(gk272) from 55 to 73%. In order to better test this hypothesis a triple mutant was constructed between lin-17(n671), lrp-2(gk272), and cam-1(gm122) (Table 1). This triple mutant displays the same P-Rvl penetrance as both the lin-17(n671); lrp-2(gk272) and lin-17(n671); cam-1(gm122) double mutants confirming that lrp-2 functions in the same pathway as cam-1.
Methods
Request a detailed protocolStrains were constructed by standard procedures (Ferguson and Horvitz, 1985).
Reagents
Strains:
N2
MT1306: lin-17(n671) (Ferguson and Horvitz, 1985)
PS4818: lin-18(e620)
PS3976: lin-17(n671)/ hT2 [qIs48 (myo-2::gfp etc.)]; +/ hT2 [qIs48]; lin-18(e620)
PS5401: egl-20(hu120)
PS5530: lin-17(n671); egl-20(hu120)
NG2615: cam-1(gm122)
RB1125: vang-1(ok1142)
VC543: lrp-2(gk272)
VC621: lrp-2(gk292)
PS4809: lin-17(n671); cam-1(gm122)
PS5651: lin-17(n671); vang-1(ok1142)
lin-17(n671); lin-18(e620) animal awere segregated from PS3976.
The lin-17(n671); lrp-2(gk292) double mutant was constructed by crossing lrp-2(gk292) males with strain MT1488: lin-17(n671); unc-13(e1091) hermaphrodites.
The lin-17(n671); lrp-2(gk272) double mutant constructed by crossing lrp-2(gk272) males with strain MT1488: lin-17(n671); unc-13(e1091) hermaphrodites.
lin-17(n671); lrp-2(gk272); cam-1(gm122) Triple mutant constructed by crossing lin-17(n671); lrp-2(gk272) hermaphrodites with cam-1(gm122) and sequencing for homozygous triples.
PS6581: qIs95[pSYS-1::VENUS::SYS-1 with pttx-3::dsRed as coninjection marker];
PS6576: lin-18(e620) sem-5(n1779)
PS6580: qIs95; sem-5(n1779)
PS6579: Pegl-17::CWN-1::GFP (unnamed construct) in cwn-1(ok546); lin-18(e620)
References
Funding
Howard Hughes Medical Institute, with whom PWS was an Investigator. The National Institute of Neurological Disorders and Stroke of the National Institutes of Health under award number 1F32NS098658-01A1 awarded to PJM.
Reviewed By
Hitoshi SawaHistory
Received: July 29, 2019Accepted: August 6, 2019
Published: August 27, 2019
Copyright
© 2019 by the authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0) License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.Citation
Minor, PJ; Sternberg, PW (2019). C. elegans LRP-2 functions in vulval precursor cell polarity. microPublication Biology. 10.17912/micropub.biology.000152.Download: RIS BibTeX
