
Description
The DAF-7/TGF-β signaling pathway plays a developmental role in the dauer decision; mutants in this signaling pathway have temperature-sensitive constitutive dauer phenotypes (Golden and Riddle 1984, Thomaset al. 1993). Additionally, the DAF-7/TGF-β signaling pathway acts in later developmental stages as an environmental sensor to regulate several aspects of metabolism and behavior in response to environmental cues (Greer et al. 2008, Guimenny and Savage-Dunn 2013, Meisel et al. 2014, Fletcher and Kim 2017). Recently it was shown that the DAF-7/TGF-β signaling pathway is required for normal regulation of levels of the glutamate receptor GLR-1 (McGehee et al. 2015). The effect of the DAF-7/TGF-β signaling pathway on GLR-1 was determined both directly using GFP-tagged GLR-1 protein (GLR-1::GFP) and indirectly by measuring the frequency of spontaneous reversals, a behavior that is controlled by signaling through GLR-1 (Zheng et al. 1999). The daf-7(e1372) allele is described as a temperature-sensitive allele (Riddle et al. 1997). When this strain is grown at 15°C worms progress normally through development, and when the growth temperature is shifted to 25°C worms enter the dauer stage (Golden and Riddle 1984). Experiments investigating the effect of the DAF-7/TGF-β signaling pathway on GLR-1 were conducted by growing animals at 15°C, followed by growth at 25°C for several hours prior to performing experiments (McGehee et al. 2015). This method of growing the worms at a low temperature and then shifting to a higher temperature is frequently used in studies using DAF-7/TGF-β signaling pathway mutants. However, some of the phenotypes associated with these mutants are not temperature sensitive, for example egg laying (Trent et al. 1983), decreased pumping rate and increased fat deposition (Greer et al. 2008). These observations suggest that it is the dauer phenotype that is temperature sensitive and not necessarily the daf-7(e1372) allele.
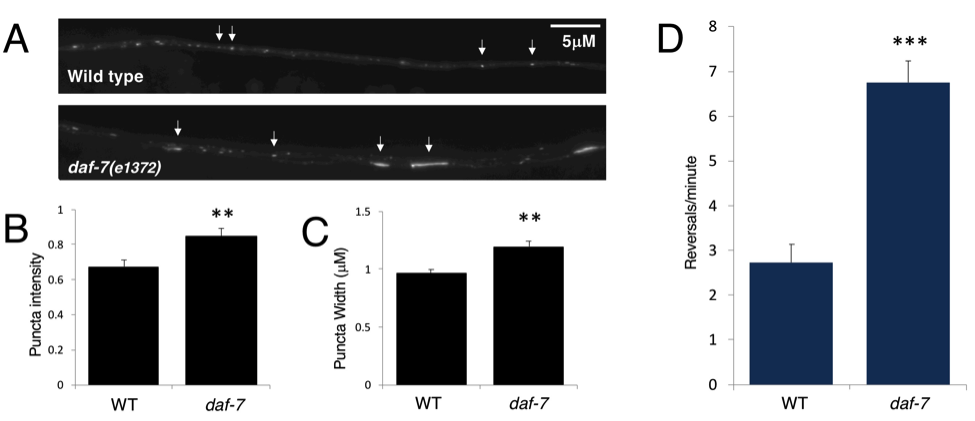
To test whether the DAF-7/TGF-β-dependent regulation of GLR-1 is temperature sensitive, animals were grown at 15°C and young adults were examined directly from 15°C cultivation (i.e. without a prior temperature shift to 25°C). Quantitative fluorescence microscopy and spontaneous reversals behavioral assays were performed as previously described (McGehee et al. 2015). The ventral nerve cord (VNC) of young adults expressing a GLR-1::GFP transgene (nuIs24 (Rongo et al. 1998)) was imaged (WT n=27, daf-7(e1372) n=27) (Figure 1A). Quanitification of images was done using MetaMorph software to generate linescans of the VNC which were analyzed in IgorPro using custom written software (Burbea et al. 2002) to quantify all of the puncta for each worm. There is variability in the puncta widths and intensities that are measured within each image, as can be seen in Figure 1A, so the average intensity and width for all of the puncta in each image was used for comparisons between the genotypes. There was a significant increase in both the intensity (p=0.002) (Figure 1B) and width (p=0.008) (Figure 1C) of GLR-1::GFP puncta in the daf-7(e1372) mutants as compared to wild type, indicating that there is more GLR-1::GFP in the daf-7(e1372) mutants. Similarly, the rate of spontaneous reversals is increased in daf-7(e1372) mutants maintained at 15°C as compared to wild type maintained at 15°C (n=11 of each genotype, p=0.000003). These results show that the previously reported GLR-1 phenotypes, namely the increased GLR-1 levels and spontaneous reversals phenotypes, are not temperature sensitive. Although it is possible that there is a secondary mutation in the daf-7(e1372) strain that causes the observed GLR-1 phenotypes, this is unlikely because the GLR-1 phenotypes are identical to those observed in mutants in other components of the DAF-7/TGF-b signaling pathway (McGehee et al. 2015). However, since transgenic rescue experiments and/or testing of a second daf-7 allele have not been performed, the presence of a secondary mutation cannot be formally ruled out. The presence of the GLR-1 phenotypes in the daf-7(e1372) strain at low temperature (15°C) is important to note as it supports the idea that it is the dauer phenotype that is temperature-sensitive and not necessarily the daf-7(e1372) allele. One implication of this finding is that it is reasonable to study the DAF-7/TGF-β-dependent control of the GLR-1 protein using the daf-7(e1372)strain in animals that have been reared at 15°C and not exposed to a temperature shift to 25°C. Additionally, caution should be used when interpreting phenotypes in this strain, and the temperature dependence of novel phenotypes should be determined.
Reagents
| Strain name | Genotype | Available from the CGC |
| N2 | Yes | |
| CB1372 | daf-7(e1372)III | Yes |
| KP1147 | nuIs24(Pglr-1::glr-1::gfp)IV | No (Rongo et al. 1998) |
| KP3079 | daf-7(e1372)III; nuIs24(Pglr-1:glr-1::gfp)IV | No (McGehee et al. 2015) |
Acknowledgments
Thank you to the following for providing strains: Joshua Kaplan, Peter Juo and the CaenorhabditisGenetics Center (CGC) (Supported by an NIH Office of Infrastructure Support Program Grant P40 OD010440). Thanks to Peter Juo for helpful discussions and microscope access.
References
Funding
Start-up funding from Suffolk University.
Reviewed By
AnonymousHistory
Received: August 7, 2019Accepted: August 19, 2019
Published: August 26, 2019
Copyright
© 2019 by the authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0) License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.Citation
McGehee, A (2019). The GLR-1 phenotypes of the daf-7(e1372) allele are not temperature sensitive. microPublication Biology. 10.17912/micropub.biology.000158.Download: RIS BibTeX
