
Description
This integrations article considers data from Kotak et al. 2019 and Trachtman et al. 2019. Our interest is in understanding the place of chromatin modifiers in developmental control pathways, working in concert with or in parallel to established transcriptional factor networks, hormonal signaling, etc.
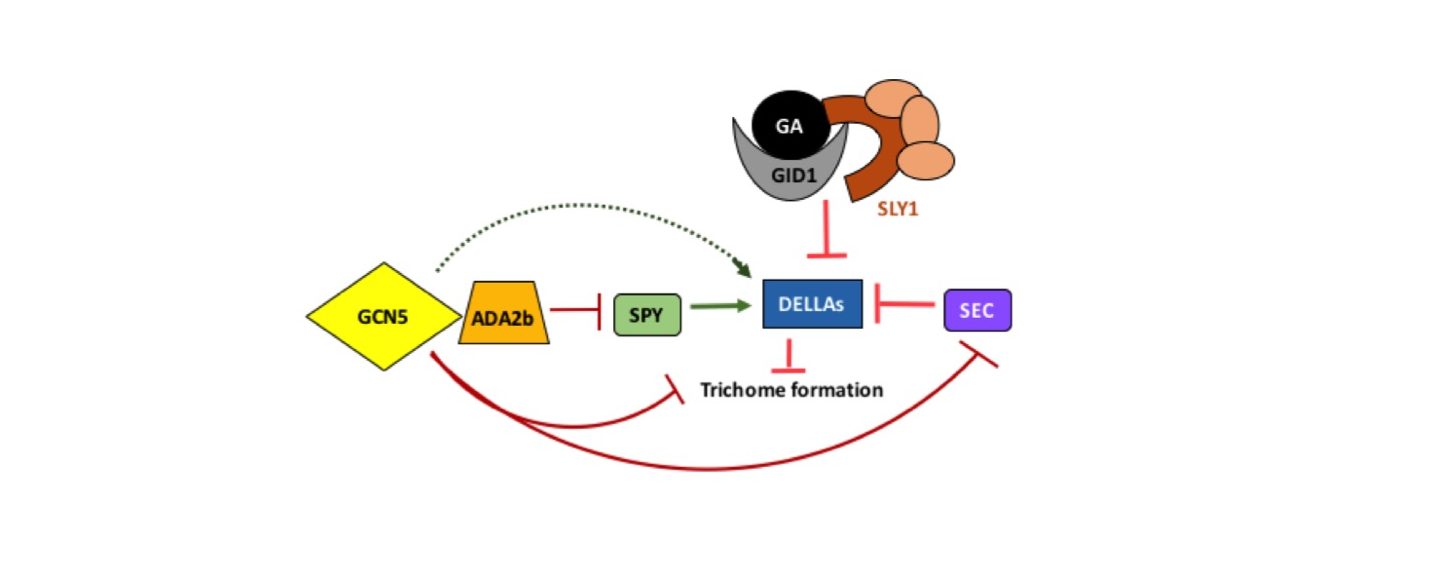
Previously we have described how the histone acetyltransferase GCN5 and its associated transcriptional coactivator ADA2b affect trichome morphogenesis (Kotak et al. 2018). We have now shown that these epigenetic factors also play a role in specification of trichome cell fate, as evidenced by an increase in trichome number and/or density in disruption mutant backgrounds (Kotak et al. 2019). Wang et al. 2019 also recently showed an increase in trichome density on the first pair of true leaves in several gcn5 mutant backgrounds. Therefore, GCN5 and ADA2b act to limit trichome initiation (Fig. 1) from a field of epidermal leaf cells, a well-described developmental process involving many genes (Pesch and Hulskamp 2009)
In addition, a number of phytohormones affect trichome development (reviewed in Fambrini and Pugliesi 2019). We focused on pathways connected to gibberellin (GA) signaling, which has been shown to stimulate trichome development in Arabidopsis (Chien and Sussex, 1996). Mutations in two members of the DELLA repressor protein family, RGA and GAI, can rescue the glaborous phenotype of ga1-3 mutants, indicating a role for these proteins in repressing trichome formation (Dill and Sun, 2001). The DELLAs themselves are negatively regulated by GA in concert with SLY1, which induces degradation via the proteasome pathway (Dill 2004) and by SEC, which has been shown to covalently modify RGA (Zentella et al. 2016). SPY also covalently modifies the DELLAs, in a way that promotes their activity (Zentella et al. 2017, Fig. 1).
With disruption of GCN5 or ADA2b, GAI expression in rosette leaves is slightly decreased while there is no detectable change in RGA expression (Trachtman et al. 2019; Vlachonasios et al. 2003). Decreased expression of a DELLA repressor would be consistent with the increased number and density of rosette leaf trichomes observed in gcn5 and ada2b disruption mutant backgrounds (Kotak et al.2019, Wang et al. 2019, Fig. 1). However, given the modest observed effects on expression of GAI only, it seems unlikely that this change alone fully explains GCN5’s and ADA2b’s roles in trichome initiation. Gan et al. (2007) have reported that GAI also has a role in limiting trichome branching so the effects on GAI may also relate to trichome morphogenesis.
We also show that disruption of GCN5 or ADA2b leads to an increased expression of SEC (Trachtman et al. 2019, Fig. 1). This effect is consistent with increased trichome number in gcn5 and ada2b disruption mutants. Our data suggest a role for ADA2b in limiting expression of SPY, which would be expected to decrease trichome number. This transcriptional effect on SPY may relate to the decrease in absolute number of trichomes observed only in the second true leaf in an ada2b-1 mutant background (Kotak et al. 2019). This finding could be further explored by conducting expression analysis in the first and second true leaves separately. It should also be noted that since qRT-PCR experiments do not directly measure functional protein levels, there could be translational or post-translational effects to consider.
While data from gcn5-1 and gcn5-6 is generally consistent, somewhat different effects (as seen with SLY1; Trachtman et al. 2019) may expected due to the nature of these lesions in the GCN5 locus and/or the genetic backgrounds (Ws vs Col ecotypes, respectively). It is also important to note that we examined mature rosette leaves, as it is technically difficult to isolate trichomes for this analysis, especially from the mutant plants. This may result in variability seen in some experiments (e.g. SEC in gcn5-1; Trachtman et al. 2019).
This work places epigenetic players in a developmental pathway alongside other transcriptional regulators. To uncover more details about how these factors act and interact, chromatin immunoprecipitation could be used to assess changes in histone acetylation state or potentially GCN5 and ADA2b binding at the GAI, SEC, and SPY promoter, to determine if these loci are direct targets of these chromatin modifiers.
Acknowledgments
We thank Dr. Konstantinos Vlachonasios (University of Thessaloniki, Greece) for his reading of this manuscript.
References
Funding
This work was supported by Muhlenberg College.
Reviewed By
Neil OlszewskiHistory
Received: September 12, 2019Accepted: October 7, 2019
Published: October 17, 2019
Copyright
© 2019 by the authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0) License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.Citation
Hark, AT; McCain1, ER (2019). The histone acetyltransferase GCN5 and the transcriptional coactivator ADA2b affect trichome initiation in Arabidopsis thaliana. microPublication Biology. 10.17912/micropub.biology.000176.Download: RIS BibTeX
