Department of Chemistry and Biochemistry, Old Dominion University, Norfolk, VA 23529
Current address: Department of Biochemistry and Biophysics, Brandeis University, Waltham, MA 02454
Description
Protein arginine methylation is a post-translational modification that contributes to the regulation of many cellular processes, including DNA damage repair, transcriptional regulation and protein-protein interaction (Bedford and Richard 2005). In 2003, a large-scale proteomics analysis identified a number of G protein-coupled receptors (GPCRs) as potentially being arginine methylated (Boisvert et al. 2003). However, the effect of methylation on these receptors was not examined. Functional evidence that protein arginine methylation regulates GPCR signaling was first reported for the D2-like dopamine GPCR family (the human D2 and C. elegans DOP-3 receptors) (Likhite et al. 2015), and later for the C. elegans SER-2 tyramine receptor (Bowitch et al. 2018). Protein arginine methyltransferase 5 (PRMT5) methylated a peptide fragment corresponding to a region of the third intracellular loop (3rd ICL) of each of these receptors in vitro, while changing the conserved arginines in this region to alanine reduced receptor methylation (Likhite et al.2015; Bowitch et al. 2018). In cell culture, mutating the conserved putative methylation motif of human D2 decreased signaling via the receptor, leading to a decrease in D2-mediated inhibition of cAMP (Likhite et al. 2015). This, combined with behavioral studies in C. elegans, suggested that protein arginine methylation promotes D2-like dopamine receptor signaling (Likhite et al. 2015). However, the mechanism by which it does so is unknown.
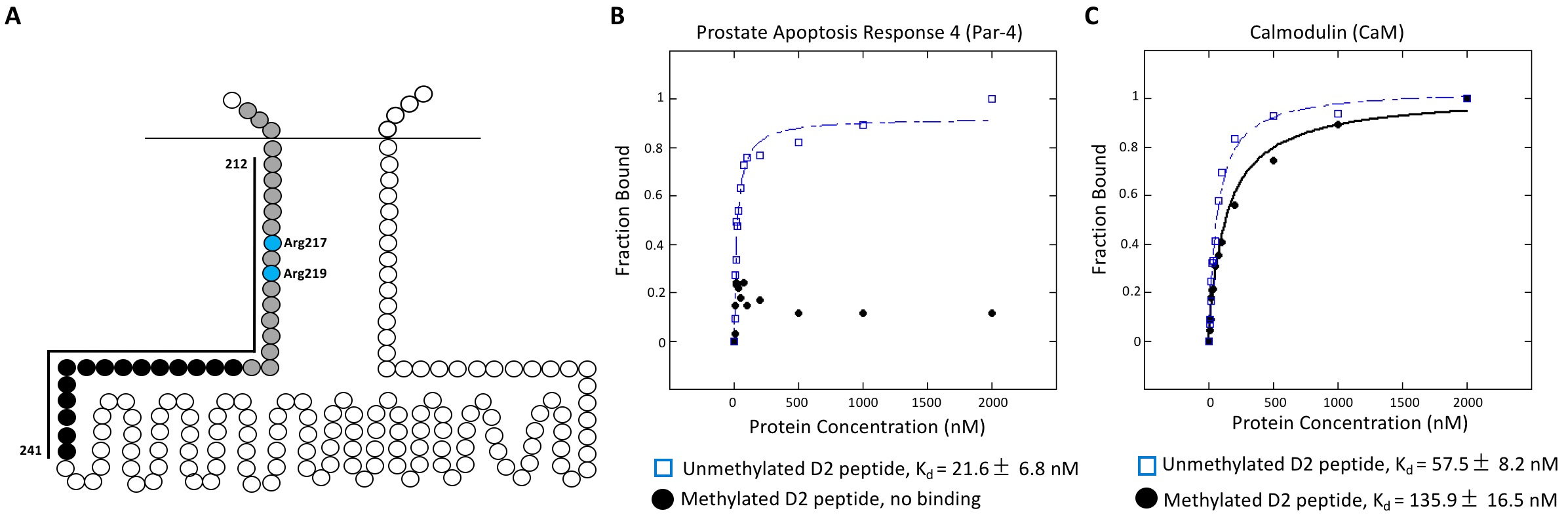
The two arginine residues (Arg217 and Arg219) that we identified as methylation targets within the amino-terminal region of the 3rd ICL of human D2 (Likhite et al. 2015) lie within the overlapping binding sites of Par-4 [prostate apoptosis response 4 protein, also known as pro-apoptotic WT1 regulator (PAWR)] and CaM (calmodulin) (Bofill-Cardona et al. 2000; Park et al. 2005) (Figure 1A). The two proteins compete for binding to D2, with CaM displacing Par-4 in a calcium-dependent manner (Park et al. 2005). Whereas Par-4 binding facilitated D2 inhibition of adenylyl cyclase (Park et al. 2005), CaM binding to D2N (a peptide corresponding to the first 19 amino acids of D2’s third intracellular loop) selectively blocked G protein activation (Bofill-Cardona et al. 2000). The opposing activities for Par-4 and CaM are consistent with their competition for binding to D2.
We sought to determine whether arginine methylation could regulate D2 signaling by altering the binding of Par-4 and/or CaM to D2. Specifically, as arginine methylation of D2 was proposed to promote D2 signaling (Likhite et al.2015), we hypothesized that it might either enhance Par-4 binding, or diminish CaM binding, to D2. We performed anisotropy experiments using purified recombinant human Par-4 or human CaM, with fluorescently-labeled unmethylated or methylated D2 peptide (corresponding to amino acids 208-243 of human D2), to determine binding constants. Unexpectedly, although Par-4 bound to the unmethylated D2 peptide with an average Kd = 21.6 ± 6.8 nM, no binding was observed in the presence of the methylated D2 peptide (Figure 1B). This suggests that methylation of the D2 receptor may actually serve to abrogate its interaction with Par-4, perhaps to allow binding of an alternative (unidentified) promoter of D2 signaling. However, consistent with the model that arginine methylation may modulate D2 receptor signaling in part by decreasing its interaction with CaM, we found that CaM bound more strongly to the unmethylated D2 peptide (average Kd = 57.5 ± 8.2 nM) than to the methylated D2 peptide (average Kd = 135.9 ± 16.5 nM).
Combined, we have demonstrated that the methylation status of a peptide fragment of the 3rd ICL of D2 influences interaction with two known modulators of D2 receptor signaling, Par-4 and CaM, in vitro. Future cell-based experiments to examine the effect of arginine methylation on full-length D2 receptor interaction with cytoplasmic regulators will provide additional insights into the mechanism by which this post-translational modification may promote D2-mediated dopamine signaling.
Methods
Request a detailed protocolPeptide synthesis
Amino-terminal FLAG-tagged peptide corresponding to amino acids 208-243 of the third intracellular loop of the human D2 receptor was synthesized by 21st Century Biochemicals with a K-5-FAM (Carboxyfluorescein) fluorophore at the carboxy-terminal end. Methylated peptides were symmetrically dimethylated at the arginines corresponding to Arg 217 and Arg 219 of D2.
His-Par-4 purification
Amino-terminal His-tagged human Par-4 was purified as previously described (Clark et al. 2018) and dialyzed into 200 mM NaCl, 200 µM CaCl2, 50 mM Tris pH 7.5 prior to lyophilization. The protein sample was reconstituted with water for use in anisotropy experiments.
His-Calmodulin purification
Amino-terminal His-tagged human calmodulin purification was adapted from (McCluskey et al. 2007). Rosetta E. coli transformants harboring the His-Calmodulin (in pET-17b, Novagen) plasmid were grown in Luria broth (LB) containing 100 µg/mL ampicillin. Cells were grown in shaking flasks at 37°C until they reached an OD600 of 0.6. Protein expression was induced with 0.75 mM IPTG and cultures were maintained overnight with shaking at room temperature. Cells were pelleted at 6000 x g for 15 minutes, washed once with PBS, and then stored at -80°C. Cell pellets from a 1-liter culture were resuspended in 30 mL of Buffer A (50 mM Tris-HCl pH 7.5, 1 mM CaCl2, 1 mM PMSF). Cells were lysed by two rounds of French press, cleared by centrifugation at 30,000 x g for 30 minutes and then filtered. Sample was adjusted to 10 mM imidazole and then loaded onto a 2-mL bed of Ni-NTA beads (Qiagen) that had been pre-equilibrated with 10 mL of Buffer A. Beads were washed three times with 10 mL of Buffer A. Sample was then eluted with 10 mL of Buffer A that had been adjusted to 150 mM imidazole. The elution was directly applied to a 2-mL bed of Phenyl-sepharose beads (Qiagen) that had been pre-equilibrated with Buffer A. Beads were washed three times with 10 mL of buffer A, and the sample was eluted in 5 mL of buffer B (50 mM Tris-HCl pH 7.5, 5 mM EGTA), and titrated with 50 mL of 1 M CaCl2. Protein was dialyzed into 1x TBS/15% glycerol overnight and stored at -80°C.
Anisotropy
10x stock concentrations of CaM or Par-4 were made in binding buffer (200 mM NaCl, 200 µM CaCl2, 50 mM Tris pH 7.5) at concentrations ranging from 0-20 µM. A master mix was made with binding buffer and fluorescently labeled unmethylated or methylated D2 peptide to yield a final concentration of 0.025 µM. 22.5 µL of master mix was aliquoted to each tube, and then 2.5 µL of the 10x stock concentration of either Par-4 or Calmodulin was added. Solutions were mixed by pipetting up and down three times and allowed to incubate at room temperature for ten minutes. 10 µL of each sample were pipetted into two adjacent wells in a 384-well black plate (Corning 3820), and fluorescence polarization was read in a SpectraMax i3x plate reader. Kd values were determined using KaleidaGraph.
Acknowledgments
We thank Skyler Kelly and Katie Lichtenthal for help with protein expression and purification and Gerald Koudelka for help with experimental design. We also thank Neah Likhite, Shigeo Yoshinari and Christopher Jackson for preliminary experiments and data that inspired this study.
References
Funding
This work was supported by the National Institutes of Health (R21MH101386 and R01DC015758 to DMF).
Reviewed By
AnonymousHistory
Received: January 22, 2021Revision received: February 2, 2021
Accepted: February 2, 2021
Published: February 9, 2021
Copyright
© 2021 by the authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0) License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.Citation
Bowitch, A; Sahoo, A; Clark, AM; Ntangka, C; Raut, KK; Gollnick, P; Yu, MC; Pascal, SM; Walker, SE; Ferkey, DM (2021). Methylation of the D2 dopamine receptor affects binding with the human regulatory proteins Par-4 and Calmodulin. microPublication Biology. 10.17912/micropub.biology.000366.Download: RIS BibTeX
