
Abstract
Single neuron-specific drivers are important tools for visualizing neuron anatomy, manipulating neuron activity and gene rescue experiments. We report here that genomic regions upstream of the C. elegans bHLH-PAS gene hlh-34 can be used to drive gene expression exclusively in the AVH interneuron pair and not, as previously reported, the AVJ interneuron pair.
Description
bHLH-PAS transcription factors are a metazoan-specific family of transcription factors with diverse functions within and outside the nervous system (Kewley et al. 2004). Caenorhabditis elegans contains five bHLH-PAS proteins: (1) ahr-1, an ortholog of the aryl hydrocarbon receptor (Powell-Coffman et al. 1998); (2) aha-1, an ortholog of the Aryl Hydrocarbon Receptor Nuclear Translocator (ARNT), which is a common dimerization partner for many, but not all bHLH-PAS genes (Powell-Coffman et al. 1998); (3) hif-1, an ortholog of hypoxia-inducible factor HIF1alpha (Jiang et al. 2001); (4) CKY-1, which orthology prediction tools consider to be an ortholog of NPAS4, even though its bHLH domain is very degenerate (nevertheless, CKY-1 heterodimerizes, like other C. elegans bHLH-PAS proteins, with the common AHA-1 partner protein in yeast 2 hybrid assays (Grove et al. 2009)); (5) hlh-34, which Marvvel (Wang et al. 2019) and Ortholist (Shaye and Greenwald 2011) predict to be an ortholog of both the NPAS1/3 and the SIM1/2 subgroups of bHLH-PAS proteins (Yan et al. 2014). However, HLH-34 contains neither of the domains that are found, in addition to the canonical bHLH and PAS domains, in either vertebrate NPAS (PAS_11 domain) or SIM (SIM_C) proteins. HLH-34 may therefore reflect an ancestral version of both subgroups.
In this paper, we re-assess the previously reported expression pattern of the hlh-34 gene. An intriguing previous study that analyzed feeding control in C. elegans used 2.5kb of the hlh-34 promoter in a reporter gene construct to assess expression of the gene (Cunningham et al. 2012). This reporter was described to be expressed exclusively in a single neuron pair that the authors tentatively identified as the AVJ neurons (Cunningham et al. 2012). Based on rescue experiments that the authors performed with the hlh-34 promoter, feeding behavior functions were ascribed to AVJ (Cunningham et al. 2012). Moreover, again based on the expression assignment of the hlh-34 promoter, an interesting recent study implicated the AVJ neurons in cold tolerance behavior (Takagaki et al. 2020). Lastly, the neurogenin homolog ngn-1 was recently reported to function in the AVJ neurons, based on its regulation of hlh-34 reporter expression (Christensen et al. 2020).
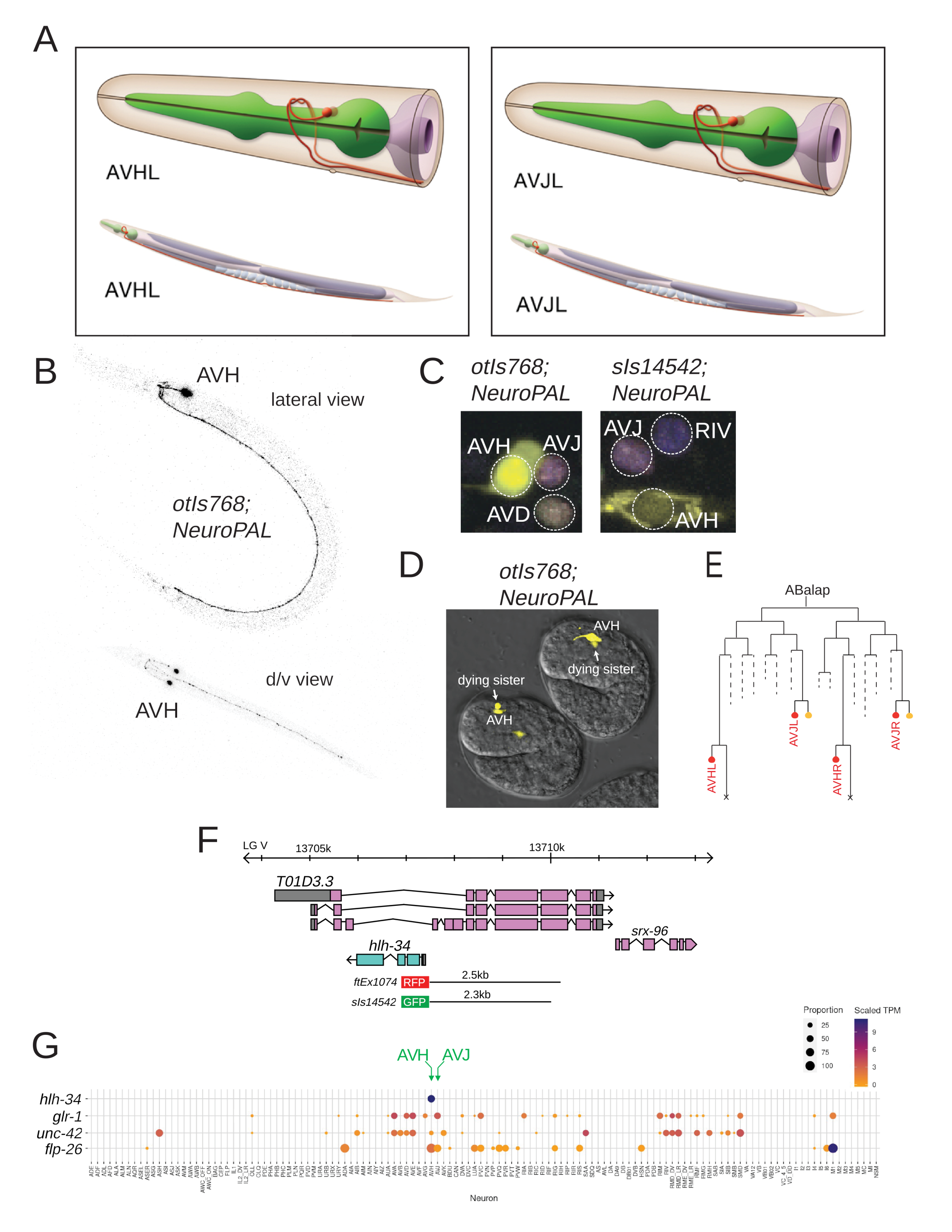
However, AVJ and its neighboring AVH neuron pair are two bilaterally symmetric neuron pairs that are notoriously difficult to distinguish based on their almost identical cell body position and neurite trajectory (White et al. 1986)(Fig.1A).
In the context of studying fate specification of the AVJ and AVH neurons (Berghoff et al. 2021), we obtained an hlh-34 promoter-based reporter line from Ian Hope, leEx1692, established in initial attempts to broadly studying gene expression (Hope et al. 1996). We found that this transgene is expressed in a single head neuron pair, extending its neurite along the ventral nerve cord (Fig.1B), consistent with this neuron being either the AVJ or AVH neuron pair. After chromosomal integration, we crossed this transgene (otIs768) with NeuroPAL, a transgene that provides a multicolor bar code for all neurons in the C. elegans nervous system, with distinct color codes for AVH and AVJ (Yemini et al. 2021). We find that the hlh-34prom::gfp signal from the otIs768 transgene overlaps with the AVH and not the AVJ signal (Fig.1C). We validated this assessment by crossing otIs768 with an AVH-expressed marker that is not in AVJ (unc-42::rfp) and an AVJ expressed marker, that is not in AVH (glr-1prom::rfp). We observed an overlap with unc-42:rfp but not glr-1prom::rfp. We also obtained another hlh-34 promoter gfp reporter from the Moerman & Baillie groups, sIs14542 (Hunt-Newbury et al. 2007) (Fig.1F). Using NeuroPAL, we confirmed sIs14542 expression in AVH, but not AVJ (Fig.1C).
We further corroborated hlh-34prom expression in AVH by observing that in the embryo, expression of otIs768 is first transiently observed in 4 cells, two of which show signs of cell death (Fig.1D); later in embryogenesis, and then during all stages of postembryonic and adult development, expression becomes restricted to 2 cells. This is consistent with expression in the bilateral AVH neuron pair, since their two sisters cells are, unlike the sisters of the AVJ neuron pair, destined to die by apotosis (Sulston et al. 1983) (Fig.1E).
Lastly, single cell RNA profiling of the entire C. elegans nervous system reveals strong and selective expression of hlh-34 transcripts exclusively in the AVH neuron (Taylor et al. 2021)(Fig.1G). The scRNA data not only confirm hlh-34 expression in AVH but also corroborates the expression of the markers that we used to distinguish AVH from AVJ (Fig.1G). Since all previously described hlh-34 reporter constructs only encapsulate parts of the 5’ region of the gene and because the gene has an unusual location in the intron of another gene (Fig.1F), the scRNA data represent an important, independent validation of the assignment of the existing reporter gene patterns to AVH and not AVJ.
The expression of the hlh-34 promoter in AVH rather than AVJ indicates that functions previously ascribed to the AVJ neuron need to be re-ascribed to the AVH neuron. This applies to the above-mentioned feeding behavior function initially ascribed to AVJ (Cunningham et al. 2012) which now has to be assigned to AVH. Similarly, the implication of AVJ in cold tolerance behavior, based on the expression overlap of a gene involved in cold tolerance, xdh-1, and hlh-34prom::rfp, and based on the rescue of the xdh-1 mutant phenotype with an hlh-34prom driver (Takagaki et al. 2020), needs to be re-assigned to the AVH neuron. Lastly, the function of the ngn-1 bHLH gene of regulating hlh-34 (Christensen et al. 2020) needs to be re-assigned to AVH.
In conclusion, the 5’ region of the hlh-34 gene is a useful tool to gain genetic access to the AVH neuron.
Reagents
The following transgenes were used:
OH16483: unc-119(ed3); otIs768[hlh-34prom::gfp, unc-119(+)]
BC15839: dpy-5(e907); sIs14542[hlh-34prom::gfp, dpy-5(+)]
OH15363: otIs669[NeuroPAL]; him-5(e1490)
Acknowledgments
We thank Kaveh Ashrafi and Ian Hope for sending us the hlh-34 reporter strains and to Wormbase for help with identifying promoter coordinates.
References
Funding
HHMI
Reviewed By
AnonymousHistory
Received: September 6, 2021Revision received: September 14, 2021
Accepted: September 14, 2021
Published: September 28, 2021
Copyright
© 2021 by the authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0) License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.Citation
Cook, SJ; Vidal, B; Hobert, O (2021). The bHLH-PAS gene hlh-34 is expressed in the AVH, not AVJ interneurons. microPublication Biology. 10.17912/micropub.biology.000467.Download: RIS BibTeX
