Duke University, Department of Biology, Regeneration Next

Description
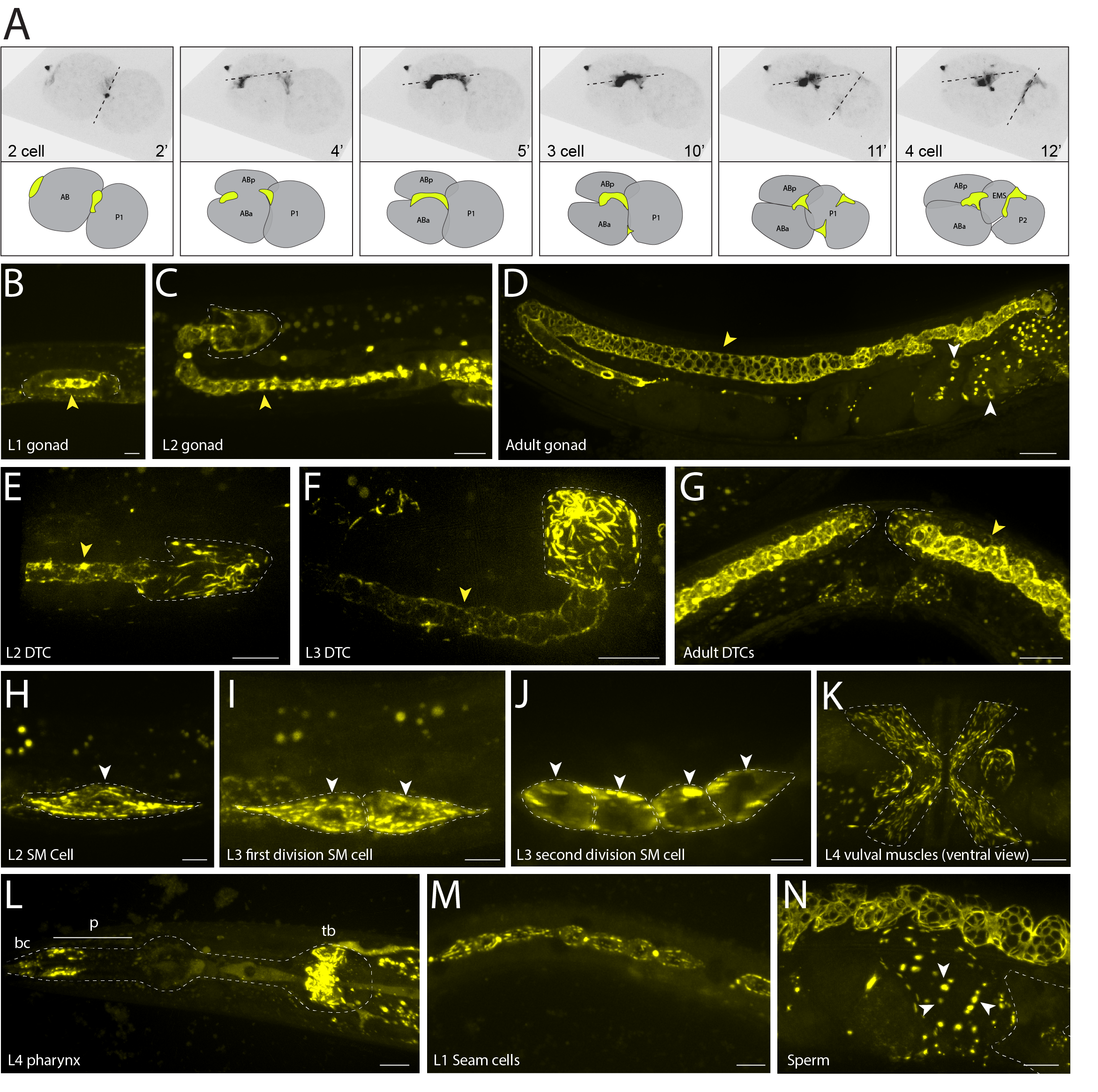
Septins are cytoskeletal proteins involved in cytokinesis, morphogenesis, and cell migration. Misregulation of septin expression occurs in neurodegenerative diseases and cancers (Angelis and Spiliotis 2016). Originally discovered in yeast, there are 13 mammalian septins and two in C. elegans, unc-59 and unc-61 (Hartwell 1971, Nguyen et al. 2000). Here, with live imaging of green fluorescent protein (GFP) fluorescence, we characterize UNC-59/Septin expression and localization throughout C. elegans development. Using CRISPR/Cas-9, we endogenously tagged C. elegans unc-59, which is most similar to human septins 1,2,4,5, and 7 (Kim et al. 2018), with GFP for visualization and live imaging using high resolution confocal microscopy. We first show the localization of UNC-59 at the cleavage furrow (previously shown with antibody staining, (Nguyen et al. 2000)) during a time lapse of cell divisions in early 2- to 4-cell stages of embryogenesis (Fig. 1A) and throughout embryogenesis (cleavage rings in older embryos in Fig. 1D). Septins are also important for gonad morphogenesis and distal tip cell (DTC) migration (Nguyen et al. 2000) where UNC-59 protein is detected throughout gonad development in the rachis (previously shown with endogenously tagged unc-59::mKate, (Priti et al. 2018)) and DTCs (Fig. 1B-G). We highlight UNC-59/Septin localization in the DTC (previously shown with a transgene, (Finger et al. 2003)) at the L2 and L3 stages where it is organized into bundles (DeMay et al. 2011) and ring structures (Figs. 1E and F). The two bilateral sex myoblast cells express UNC-59 during their posterior to anterior migration in the L2 and early L3 stage (Fig. 1H) and continue to express UNC-59 in these cells as they differentiate into vulval muscles in the late L3 to early L4 stages (Fig. 1K). Lastly, we show UNC-59/Septin expression and localization in tissue not previously reported: in the pharynx (cells of the buccal cavity, anterior procorpus, and terminal bulb) (Fig. 1L); in the seam cells, both in bundles and at the cleavage furrows, beginning in the L1 stage (Fig. 1M) and continuing throughout development and into the adult; and in sperm surrounding an embryo that has exited the spermatheca (Fig. 1N).
Reagents
CRISPR constructs were generated using a self-excising cassette (SEC) for drug (hygromycin) selection as described previously (Dickinson et al. 2015). Guide plasmids were generated using plasmid pDD122 (Peft-3::Cas9 + ttTi5605 sgRNA); Addgene 47550. SG: AAGAAACGAATGGGCGGTCTCGG. To generate unc-59(qy88[unc-59::GFP::3xflag::AID+loxP]), pDD282 (GFP-C1^SEC^3xFlag^AID) was modified (Addgene 66823). Primers to generate the homology arms (amplified from gDNA): 5’ F: tacgactcactatagggcgaattgggtaccacaactagtCGTAATGTTCATTATGAGAAT
5’ R: TGGGACAACTCCAGTGAACAATTCTTCTCCTTTACTCATGTTTCGATTAAACAATCCtAatCCtCCCATTCGTTTCTT
3’ F: AAATCAAGCGGTGGCCCGGAGGCGGCGGCGTTCGTGAATAATTCCCTCATTTTTTAAACG
3’ R: agggaacaaaagctggagctccagcggccgctttgcatgCATGTCTTTGTAATGCTGTGG
The first two codons of GFP were removed to generate a direct fusion after a recommendation from Amy Gladfelter at UNC Chapel Hill.
References
Funding
D.C. is supported by Duke URS Independent Study Grant, E.H. is supported by postdoctoral fellowship 129351-PF-16-024-01-CSM from the American Cancer Society, and D.R.S. is supported by the National Institutes of Health 5R35GM118049
Reviewed By
AnonymousHistory
Received: November 22, 2019Accepted: December 20, 2019
Published: December 20, 2019
Copyright
© 2019 by the authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0) License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.Citation
Chen, DD; Hastie, E; Sherwood, DR (2019). Endogenous expression of UNC-59/Septin in C. elegans. microPublication Biology. 10.17912/micropub.biology.000200.Download: RIS BibTeX
