
Abstract
The F-actin, spectrin, and microtubule cytoskeletons are important mediators of embryonic epidermal morphogenesis in Caenorhabditis elegans. While SMA-1/βH-spectrin is known to organize actin bundles that connect to cadherin-based adhesions, the role of microtubules in the developing epidermis is not well understood. To determine if the spectrin cytoskeleton also plays a role in organizing epidermal microtubules, we conducted feeding RNA interference of four microtubule-associated protein genes in sma-1/βH-spectrin null animals. Knockdown of apr-1, unc-33, unc-44, or cls-1 in sma-1(ru18) homozygotes did not reveal a genetic interaction; however, knockdown of hmp-2/β-catenin in sma-1(ru18) synergistically increased embryonic lethality and epidermal defects.
Description
During embryonic development, changes to cell shape, structure, and location act together to construct an organism’s overall form. Epidermal morphogenesis in Caenorhabditis elegans has been a well-studied model for these types of events as this epithelial tissue has substructures like cell-cell and cell-matrix adhesions similar to those in vertebrates (Hsiao and Chisholm 2012). Epidermal cells arise on the dorsal surface of the C. elegans embryo with different subsets of cells undergoing various morphogenetic changes to wrap the embryo in an epithelial sheet (Chisholm and Hardin 2005). During dorsal intercalation, two rows of epithelial cells interdigitate with one another to become a single row along the anterior-posterior axis (Priess and Hirsh 1986; Williams-Masson et al. 1998). Additional cells from either side of the embryo migrate towards the ventral midline, forming new cell-cell adhesions with their contralateral neighbors (Priess and Hirsh 1986; Williams-Masson et al. 1997; Costa et al. 1998). Contractions along circumferential F-actin bundles in the epidermis then change the shape of the cells to drive elongation of the animal along the anterior-posterior axis (Priess and Hirsh 1986). The spectrin cytoskeleton lies apically between the plasma membrane and circumferential F-actin bundles and is thought to be important in structurally supporting the F-actin network during these contractions (Praitis et al. 2005; Lardennois et al. 2019). Homozygous mutant animals of sma-1(ru18), a putative null allele of sma-1/βH-spectrin, are shorter than wildtype with severe disruptions in F-actin bundles normally located at the apical surface (McKeown et al. 1998; Praitis et al. 2005). Since F-actin bundles are also anchored to cadherin-based adhesions at dorsal-ventral cell boundaries, disruption of HMR-1/cadherin, HMP-2/β-catenin, or HMP-1/α-catenin also results in elongation defects and a loss of F-actin at adherens junctions, as well as embryonic lethality (Costa et al. 1998).
Microtubules are organized in a similar pattern to F-actin in the embryonic epidermis and temporally-controlled experiments have shown that they are also required for proper body elongation (Priess and Hirsh 1986; Quintin et al. 2016). It is possible that the spectrin cytoskeleton supports the structure and function of the microtubule network as it does for F-actin, but currently no studies have looked at such a connection. Since microtubules are not only essential for epidermal morphogenesis but also early cell divisions, complete disruption would make it difficult to study an interaction with spectrin. Microtubule-associated proteins (MAPs) control many of the important features of microtubules, from dynamic instability to vesicular trafficking (Alfaro-Aco and Petry 2015), and there are at least 60 conserved MAPs in C. elegans (Lacroix et al. 2014). If there is an interaction between the spectrin cytoskeleton and microtubules that supports proper epidermal morphogenesis, perhaps this can be revealed by weak disruption of specific MAPs in a sensitized, sma-1/βH-spectrin null background.
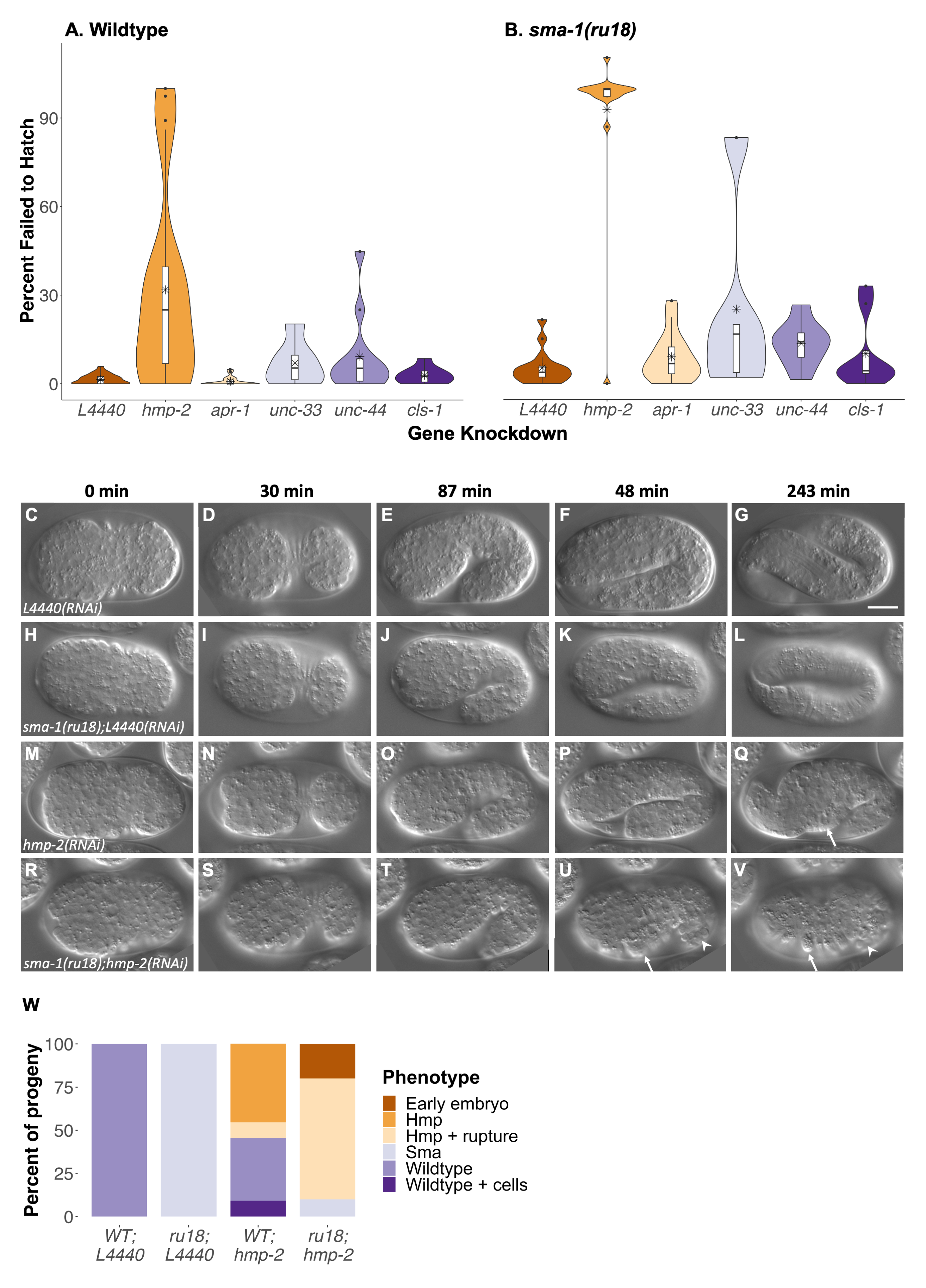
RNA interference (RNAi) by feeding is known to cause a weaker knockdown for many genes when compared to RNAi by injection (Timmons and Fire 1998). To take advantage of this, we conducted feeding RNAi of four MAP genes (apr-1, unc-33, unc-44, cls-1) in wild-type and sma-1(ru18) homozygous animals and determined the number of progeny that failed to hatch (Fig. 1A-B). In wildtype (Fig. 1A), two-way ANOVA with Tukey comparison found no significant differences in embryonic lethality between the L4440 empty vector RNAi control and any of the four MAP genes tested (apr-1, p=1.00; unc-33, p=1.00; unc-44, p=0.98; cls-1, p=1.00). Consistent with previous findings, sma-1(ru18) animals fed the L4440 control showed very low levels of embryonic lethality (Fig. 1B) similar to those found in the wild-type L4440 control (Fig. 1A; p=1.00). RNAi of the four MAP genes in sma-1(ru18) homozygous animals also showed no significant changes in embryonic lethality when compared to either the sma-1(ru18) L4440 control (apr-1, p=1.00; unc-33, p=0.48; unc-44, p=0.99; cls-1, p=1.00) or the same treatment in wildtype (apr-1, p=0.97; unc-33, p=0.83; unc-44, p=1.00; cls-1, p=1.00). It is important to note that knockdown by feeding RNAi may have been ineffective for these four MAP genes. For example, since roughly 65% of apr-1/APC homozygous null animals are embryonic lethal (Hoier et al. 2000), one would expect even weak apr-1 knockdown in wildtype to exhibit some significant lethality. At the very least the results indicate that if these MAPs play a role during epidermal morphogenesis, the sma-1(ru18) sensitized background is insufficient to reveal that role.
In contrast, RNAi of hmp-2/β-catenin in the sma-1(ru18) background did reveal an important genetic interaction during embryogenesis. As a positive control in wild-type animals, hmp-2(RNAi) resulted in an average of 31.8% embryos that failed to hatch, which was significantly different than the average 1.5% of embryos that failed to hatch in the RNAi vector control, L4440 (Fig. 1A; p<0.01). In sma-1(ru18) homozygous animals, hmp-2 knockdown resulted in even greater embryonic lethality, with 92.8% of embryos that failed to hatch compared to 5.3% in L4440 (Fig. 1B), a statistically significant increase of 61.0% over the hmp-2 knockdown in wildtype (p<0.01).
To determine the cause of the increased embryonic lethality in sma-1(ru18); hmp-2(RNAi) animals, we used time-lapse differential interference contrast (DIC) microscopy (Fig. 1C-V) and categorized the resulting phenotypes (Fig. 1W). In wild-type control embryos (Fig. 1C-G), epidermal cells properly migrated from the dorsal surface to the ventral midline (Fig. 1C-D) before animals elongated along the body axis (Fig. 1E-G). Consistent with the literature, homozygous sma-1(ru18) embryos (Fig. 1H-L) completed ventral enclosure successfully (Fig. 1H-I) but body elongation was stunted (Fig. 1J-L), with 100.0% of embryos reaching only two-fold their original length (Fig. 1L) compared to the four-fold increase seen in wild-type animals (Fig. 1G). In wild-type embryos with hmp-2 knockdown (Fig. 1M-Q), 45.5% of animals showed elongation defects (Fig. 1O-Q) similar to those of sma-1(ru18) homozygotes (Fig. 1J-L) but with additional body morphology defects (i.e., humps) (Fig. 1Q; arrow). In 9.0% of wild-type hmp-2(RNAi) embryos, animals appeared to elongate four-fold but extraneous material, possibly cells, was observed in the egg shell (Fig. 1W). In sma-1(ru18); hmp-2(RNAi) embryos, 70.0% of embryos elongated to only 1.5-fold their original body length (Fig. 1T-U) before forming humps (Fig. 1U-V; arrow), with each of these embryos exhibiting cell ruptures from the posterior after the onset of humps (Fig. 1U-V; arrowhead). In 20.0% of sma-1(ru18); hmp-2(RNAi) embryos, the landmarks of dorsal intercalation or ventral enclosure were not apparent (Fig. 1W; Extended Data), indicative of an earlier defect in embryogenesis.
In hindsight, a genetic interaction between sma-1/βH-spectrin and hmp-2/β-catenin could have been predicted based on the roles of these two genes in epidermal morphogenesis. At sites of cell-cell adhesion in the embryonic epidermis, HMP-2/β-catenin acts as a bridge between HMR-1/cadherin and the F-actin binding protein HMP-1/α-catenin (Costa et al. 1998; Maiden et al. 2013). Loss of HMP-2 therefore results in a complete loss of F-actin at the adherens junction (Costa et al. 1998), whereas sma-1(ru18) homozygous mutants maintain F-actin at adherens junctions but the circumferential actin bundles are disorganized (Praitis et al. 2005). Even though hmp-2 and sma-1 perturbation both disrupt epidermal F-actin, our expectation was that the embryonic lethality observed in hmp-2(RNAi) embryos would simply supersede the mild elongation defects in sma-1(ru18) homozygous mutants and that the percent of embryos that failed to hatch in sma-1(ru18); hmp-2(RNAi) would be similar to hmp-2(RNAi) in wild-type animals. The earlier formation of humps and the cells rupturing from the posterior in sma-1(ru18); hmp-2(RNAi) embryos, however, would indicate that the genetic interaction between sma-1 and hmp-2 is important in maintaining stable cell-cell adhesions due to the roles of each in proper F-actin attachment and organization. However, the 20.0% of embryos that appeared to have defects earlier than dorsal intercalation or ventral enclosure may be evidence of a role for these two genes in the cell shape changes occurring during gastrulation. As observed in the embryonic epidermis, SMA-1 may be required to properly organize and maintain actin bundles along the apical membrane surface in cells undergoing apical constriction, like the endoderm precursor cells (Goldstein and Nance 2020). HMP-2/β-catenin and HMP-1/α-catenin have already been shown to be required for HMR-1/cadherin to become more apically enriched at lateral contacts between these endoderm precursor cells during gastrulation (Marston et al. 2016), so disruption of both sma-1 and hmp-2 could more severely perturb the organization of actin and cadherin-based adhesions in these cells. A role for βH-spectrin in gastrulation has already been found in Drosophila ventral furrow invagination, where a loss of βH-spectrin prevents stabilization of the cell surface during the pulsatile events of apical constriction (Krueger et al. 2020). A similar but mechanically distinct ratcheting mechanism has been proposed for C. elegans SPC-1/α-spectrin in the muscle-dependent stage of embryonic elongation (i.e., past two-fold) (Lardennois et al. 2019). Further mechanistic studies will be needed to determine the potential role of SMA-1/βH-spectrin and HMP-2/β-catenin during C. elegans gastrulation.
Methods
Request a detailed protocolC. elegans strains were cultured using standard protocols (Brenner 1974). Strains used include N2 [wildtype] and AZ30 [sma-1(ru18)].
Bacterial clones for feeding RNAi were obtained from the Ahringer library (Kamath and Ahringer 2003). The hmp-2 gene insert was verified by Sanger sequencing through Eurofins Genomics using the following vector primers: GTCAGTGAGCGAGGAAGCAAC and CTCTTCGCTATTACGCCAGCTG. For knockdown by feeding RNAi, bacterial cultures were plated on NGM plates supplemented with 25 μg/mL carbenicillin, 10 μg/mL tetracycline, and 1 mM IPTG, and incubated for 3 days at room temperature to induce double-stranded RNA. L4 worms at 20 °C were fed bacteria for 48 hours and then singled to individual plates. After laying eggs for approximately 18 hours, the adults were removed and all progeny were counted via a dissecting microscope. After an additional 24 hours, the remaining number of eggs were counted. The violin plot, basic summary statistics, two-way ANOVA Type III, and the post-hoc Tukey analysis were all completed using R.
For time-lapse DIC microscopy, eggs were collected from gravid hermaphrodites after L4 worms were fed RNAi bacteria for 48 hours. Eggs were mounted on a 5% agarose slide and imaged using 1-μm slice spacing throughout the embryo at 3-minute intervals over 9 hours using a Leica DIC microscope with a 63x/1.25 NA oil HCX PL FLUOTAR objective at 20 °C with a Windows PC computer running IPLab 4.0 software and a Qimaging QICAM Fast 1394 camera. ImageJ was then used to analyze the 4D dataset.
Acknowledgments
Some strains were provided by the CGC, which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440). Ahringer library clones were kind gifts from Jeff Hardin. Special thanks to Joey Hubbard for help with the statistical analysis and Vida Praitis for critical reading of the manuscript.
References
Funding
Department of Biology, Truman State University
Reviewed By
AnonymousHistory
Received: June 22, 2021Revision received: June 29, 2021
Accepted: June 30, 2021
Published: July 13, 2021
Copyright
© 2021 by the authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0) License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.Citation
Wieberg, S; Euwer, H; Gerst, A; Maiden, SL (2021). Synergistic effects of hmp-2/β-catenin and sma-1/βH-spectrin on epidermal morphogenesis in Caenorhabditis elegans. microPublication Biology. 10.17912/micropub.biology.000417.Download: RIS BibTeX
